INFECTIOUS PANCREATIC NECROSIS VIRUS (IPNV) AS RISK FACTOR IN THE DEVELOPMENT OF HEART AND SCELETAL MUSCLE INFLAMMATION (HSMI) IN ATLANTIC SALMON (SALMO SALAR L.).
Introduction
Heart and skeletal muscle inflammation (HSMI) is a viral disease which produce severe losses in Atlantic salmon farming due to mortality, reduced growth and feed conversion and quality of meat. Causative agent Piscine reovirus (PRV) is found both in wild and farmed Atlantic salmon. Most disease outbreaks are registered after transfer to seawater. No vaccines are available and little is known about the host-pathogen interaction and what factors that can affect disease development. IPNV is a prevalent pathogen in Norwegian farmed Atlantic salmon. The virus causes substantial mortality in juvenile Atlantic salmon and smolts when transferred to seawater and persistent infections in surviving fish after disease outbreaks. IPN registered the first weeks after seawater transfer is most likely due to recurrence of an IPN virus carrier infection established during the freshwater period. HSMI often follows after IPN outbreaks, and we therefore wanted to study if IPN virus infections affect the development of HSMI.
Materials and methods
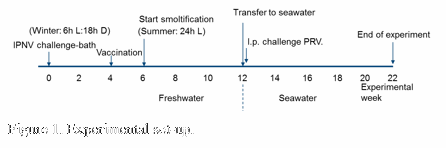
Atlantic salmon parr, mean weight 20 gr, were challenged by bath with IPN virus in freshwater (see figure 1) to establish an experimental carrier condition. All salmon in the farming industry are vaccinated before transfer to sea. Four weeks after infection we therefore vaccinated half of the fish with a commercial multivalent vaccine containing bacterial and IPNV antigens while the other half were left unvaccinated. At transfer to seawater half of the vaccinated and unvaccinated fish were challenged by intraperitoneal (i.p.) injection of heart homogenate containing high levels of PRV. Organ samples for IPNV and PRV detection were harvested regularly throughout the whole experiment. Three, 7 and 10 weeks after PRV challenge heart samples were harvested for verification of HSMI by histopathological analyses.
Results
No mortalities were registered after bath challenge with IPNV or after i.p. challenge with PRV. Head kidney samples from IPNV challenged fish were tested by titration in cell culture and by qPCR to follow the development of IPNV titers throughout the 22 week experiment. Infective virus could be detected at all time-points, with the highest levels and frequences 3 weeks after challenge. Viral transcripts were found at a higher frequency than infective virus. The virus could be detected in both vaccinated and unvaccinated fish. Levels of IPN virus were not significantly affected by the PRV infection, but there was a tendency of lower levels of IPNV transcripts in PRV infected fish. No re-activation of the carrier IPN virus infection was observed after transfer to seawater. Of the PRV challenged fish tested, 59/60 was diagnosed with HSMI 7 weeks after PRV challenge by histological analyses of heart samples. PRV levels were higher 7 weeks after infection compared to week 3 and 10. No differences in heart pathology could be registered between IPNV challenged and unchallenged fish at any time-points, however significantly higher amounts of PRV transcripts were detected at week 7 and 10 in vaccinated compared to unvaccinated fish.
Discussion and conclusion
All groups were infected with IPNV after bath challenge in freshwater and at group level a long-lasting carrier state was established. No reactivation of the virus was observed after seawater transfer, as has earlier been demonstrated in similar experimental set-ups (Stangeland et al 1996, Johansen et al 2001, 2009, Julin et al submitted). The PRV infection and the following development of HSMI did not significantly affect the levels of IPNV. The same has earlier been demonstrated in Infectious salmon anemia virus (ISAV) infected A. salmon carrying IPNV (Melby and Falk, 1995). Maximum PRV levels were detected at week 7 post infection, corresponding to the time-point were 59/60 fish were diagnosed with the disease. HSMI developed in both IPNV carriers and non-carriers. No unspecific protection against HSMI was seen in the vaccinated groups, as has earlier been demonstrated for IPN (Eggset et al 1997, Johansen et al 2001). The reason for, and possible consequences of, the elevated level of PRV in vaccinated fish compared unvaccinated fish remains to be elucidated.
References
Eggset et al 1997, Aquaculture 158: 179-191
Johansen and Sommer 2001, DAO 47: 109-117.
Johanssen et al 2009, Aquaculture 290: 9-14.
Melby and Falk, 1995, J Fish Dis 18: 579-586.
Stangeland et al 1996, J Fish Dis 21: 193-204.