THE EFFECT OF DIFFERENT LIVE FEED ORGANISMS ON LARVAL BALLAN WRASSE Labrus bergylta
Introduction:
The use of cleaner fish is one important tool in the tool-box to combat the sea lice problem in salmon farming. The major proportion of wrasse used as cleaner fish by the Norwegian salmon industry is coming from capture-based activities, depleting wild stocks. Amongst the four wrasse species used, ballan wrasse (Labrus bergylta) is the only species which is also cultivated, though only about 30% of the approx. 3 million fish used per year is produced in aquaculture (Fiskeridirektoratet 2021). The major obstacle in wrasse cultivation preventing large scale production is the difficult first feeding phase and a lack of functional feeding protocols.
Like most marine fish larvae ballan wrasse do not accept inert diets as feed. The current solution to this problem is the use of rotifers as the first feed with a transition to artemia nauplii after about 2 weeks. The initiation of formulated diets usually takes place around 4 weeks into the life of a ballan wrasse larvae. Feeding regimes based on rotifers and artemia usually produce mixed results and are way from being optimal for the larvae (Øie, Galloway et al. 2017), and there is good evidence that effects implanted into any animal during early life history have lasting effects (Winick and Noble 1966).
Material & Methods:
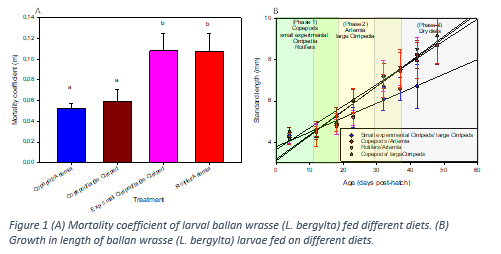
In a 48-day start feeding experiment with ballan wrasse, we studied the feasibility of replacing (Phase 1) rotifers by an experimental cirriped diet or copepod nauplii (Acartia tonsa), and (Phase 2) replacing artemia by nauplii of the cirriped Semibalanus balanoides. During phase 3 all treatments received the same commercially available formulated diets. Four different treatments were studied: (1) Control: Rotifers-Artemia-Dry diets; (2) Copepods-Artemia-Dry diets; (3) Copepods-Large cirripedia-Dry diets; and (4) Small experimental cirriped diet- Large cirripeds-Dry diets. We sampled at each feed transition for a suite of different response variables such as growth, morphometry, gene expression, lipidomics, histology, and microbiology.
Results:
We found significant differences in mortality rates. Larvae started with copepods died at significantly lower rates than larvae started on either rotifers or small experimental cirripeds (Figure 1 A). The experimental cirripede diet comprised a mix of small and large cirripede nauplii at a ratio of 1:1, whereof the larger nauplii were too large for the larvae to ingest which affected the performance of this group.
This pattern was also reflected in early growth (Figure 1 B), copepods fed larvae grew faster than the other two groups. However, once the rotifer-started larvae were fed artemia, they picked up growth rates which were then comparable to larvae receiving copepods in the beginning. There was no pronounced difference between copepod started larvae feeding on artemia or large cirripeds during phase 2 of the experiment.
Gene expression studies at day 42 post-hatch revealed pronounced differences between the rotifer started group and the other three treatments which received natural, unenriched diets. Here, genes involved in fatty acid elongation, terpenoid biosynthesis and purine/pyrimidin metabolism were significantly higher expressed in the rotifer started group. Pronounced differences were also observed in amino acid metabolism. Within the three unenriched groups differences in gene expression were very small. Lipidomics revealed that a group of phosphatidylethanolamines (PE) correlated negative with larval growth rates, while a group of triacylglycerids (TAG) correlated positive with growth. Contrary, a group of monoacylglycerids (MAG) correlated positive with mortality rates.
Discussion & Conclusion:
The first diets in the life of the of the larvae clearly had effects on growth and mortality, but more important, clear effects of the first feed were still visible in both, gene expression pattern and lipidomics data at the end of the experiment. At this time, the larvae received other live feed organisms and the same formulated diets for three weeks. This clearly points towards a nutritional programming in larval fish, as described e.g. in mammals (Hou and Fuiman 2019). This concept opens unique opportunities to not only increase our knowledge on the importance of early life history nutrition in larval fish, but also to use this knowledge to program metabolic pathways early in the life of a fish to increase fish welfare throughout the whole lifespan.
References:
Fiskeridirektoratet. (2021). "Use of cleaner fish in the production of Atlantic salmon and Rainbow trout." from https://www.fiskeridir.no/Akvakultur/Tall-og-analyse/Akvakulturstatistikk-tidsserier/Rensefisk.
Hou, Z. and L. A. Fuiman (2019). "Nutritional programming in fishes: insights from mammalian studies." Reviews in Fish Biology and Fisheries.
Winick, M. and A. Noble (1966). "Cellular response in rats during malnutrition at various ages." The Journal of nutrition 89(3): 300-306.
Øie, G., T. et al (2017). "Effect of cultivated copepods (Acartia tonsa) in first-feeding of Atlantic cod (Gadus morhua) and ballan wrasse (Labrus bergylta) larvae." Aquaculture Nutrition 23(1): 3-17.