DIETARY EFFECTS ON THE REPRODUCTIVE PERFORMANCE OF THE SEA URCHIN Tripneustes gratilla
Introduction
Tripneustes gratilla has been identified as a commercially viable species for aquaculture production in South Africa, with the potential for inclusion in integrated multi-trophic aquaculture (IMTA) systems. However, these broadcast spawning animals often display differential parental contributions within aquaculture environments. This reproductive strategy can reduce genetic variation through a bottleneck effect, subsequently resulting in a poor response to artificial selection and poor production output in cultured populations (Grant et al., 2017). Additionally, feed-type and quality, as well as the feeding regimes used for broodstock conditioning, will impact feed intake, digestion and eventual nutrient availability, affecting gametogenesis and reproductive performance of these animals (Azad et al., 2011). This study assessed various biological and genetic factors for T. gratilla, as well as different feeding regimes that could affect reproductive competition, larval growth and juvenile performance after implementing a factorial breeding design.
Materials and Methods
First-generation (F1) broodstock animals (n = 32) were conditioned on 4 diets [formulated feed (20% Ulva inclusion (Cyrus et al., 2014)), kelp (Ecklonia maxima), green seaweed (Ulva rigida) and mixture of the three diets] for approximately four months. Spawning was induced by injecting 0.5M KCl. Sperm or eggs were collected for quantification and subsequent morphological assessments, as well as for the quantification of egg lipids, proteins, carbohydrates and fatty acid profile assessments using gas chromatography flame ionisation detection (GC-FID). Remaining gametes were used in a factorial breeding design, where a standardised number of sperm and eggs of each individual were combined with that of other individuals fed the same broodstock conditioning diet. After spawning, broodstock animals were dissected, gonad weight and colour assessed and gonads processed for routine paraffin histology to evaluate gonad maturity.
Larvae were reared in 130L conical tanks. Larval morphology and survival were assessed for 20 days. Larvae from the kelp and mixed diet fed broodstock survived for the full duration of larval rearing and were transferred to 87L settlement containers, pre-coated with Ulvella lens. Approximately 3 months post-settlement, DNA was extracted and 10 species-specific microsatellite markers (Carlton and Lippé 2007; Wainwright et al., 2012) were amplified across 16 F1 broodstock (8 fed kelp and 8 fed mixed diet) and a total of 364 second-generation (F2) offspring. Genetic diversity of F1 broodstock relative to the F2 offspring, parental contribution through parentage assignment and heritability (h2) estimates for growth were evaluated.
Results and Discussion
Across broodstock, significant differences (P < 0.05; ANOVA) in gonad and egg colour suggest a formulated feed (20U) should not be fed in isolation for reproductive purposes. Dietary carotenoids (red, orange and yellow pigments) incorporated in sea urchin gonads and eggs could have downstream effects on urchin health and reproduction; via pro-vitamin A, photoprotective, and immunity-related roles, resulting in improved hatching and larval survival. Excess linoleic acid (C18:2n6) was observed in samples from animals fed a formulated feed. Although these fatty acids may elicit inflammatory response in urchins, there are potential benefits to including a formulated feed in conjunction with natural feeds in a mixed feeding regime. The egg fatty acid profile associated with the mixed diet clustered separately (principal component analysis) from the singular feeds, possibly as a result of the interactions between the combination of compounds obtained from the individual diets.
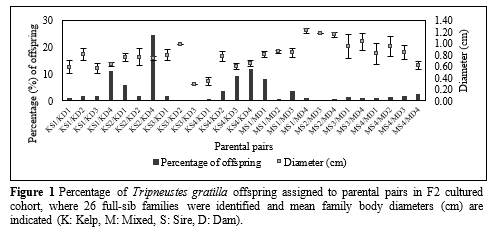
Larvae from broodstock fed kelp and the mixed diet displayed similar growth rates, with larvae from broodstock fed a mixed diet displaying a greater extent of phenotypic plasticity. Genetic diversity analyses showed no statistically significant (P > 0.05) differences between F1 broodstock animals and their F2 offspring, with an average allelic richness (AR) of 5.31 ± 0.52 and 5.18 ± 0.42, respectively. A total of 26 out of 32 possible parent pairs contributed to the F2 generation (Fig. 1) and juveniles assigning to broodstock fed a mixed diet were significantly (P < 0.05) larger (average offspring size of 0.94 ± 0.10 cm) than broodstock fed kelp (average offspring size of 0.66 ± 0.07 cm). However, a larger number of offspring (79.65%) were assigned to kelp fed broodstock (Fig. 1). An assessment of offspring phenotypic performance showed low heritability estimates (h2 = 0.050 ± 0.058) for body diameter, suggesting that additive genetic effects play limited roles in this trait.
Conclusion
Aquaculture establishments could take advantage of the maternal provisioning strategy of sea urchins to benefit future commercial production. Animals fed the mixed feeding regime outperformed the other feeds across various measurements, with relatively equal parental contributions to offspring. The varied nutrient content of the included feeds, carotenoid content, diverse array of essential amino acids and fatty acids supplied by the respective singular feeds, as well as improved digestibility through the enzymatic activity of the bacterial communities of the natural feeds, could have contributed to the reproductive success of the mixed diet fed broodstock. This study also showed that the implementation of a factorial breeding design maximises genetic diversity in subsequent generations by negating unequal parental contributions to some extent.
References
Azad, A.K., Pearce, C.M., McKinley, R.S., 2011. Effects of diet and temperature on ingestion, absorption, assimilation, gonad yield, and gonad quality of the purple sea urchin (Strongylocentrotus purpuratus). Aquaculture 317: 187–196.
Carlton, D.B., Lippé, C., 2007. Eleven new microsatellite markers for the tropical sea urchin Tripneustes gratilla and cross-amplification in Tripneustes ventricosa. Molecular Ecology Resources 7: 1002–1004.
Cyrus, M.D., Bolton, J.J., de Wet, L., Macey, B.M., 2014. The development of a formulated feed containing Ulva (Chlorophyta) to promote rapid growth and enhanced production of high quality roe in the sea urchin Tripneustes gratilla. Aquaculture Research 45: 159–176.
Grant, W.S., Jasper, J., Bekkevold, D., Adkison, M., 2017. Responsible genetic approach to stock restoration, sea ranching and stock enhancement of marine fishes and invertebrates. Reviews in Fish Biology and Fisheries 27: 615–649.
Wainwright, B.J., Arlyza, I.S., Karl, S.A., 2012. Isolation and characterization of nineteen microsatellite loci for the collector sea urchin, Tripneustes gratilla. Conservation Genetic Resources 4: 963–965.