POPULATION STRUCTURE AND GENETIC VARIABILITY BASED ON GENOMIC INFORMATION IN GILTHEAD SEABREAM AND EUROPEAN SEABASS POPULATIONS
Introduction
Gilthead sea bream (Sparus aurata) and European seabass (Dicentrarchus labrax ) are the most important marine farmed fish in the Mediterranean . In recent years, different companies have initiated breeding programmes and the proportion of genetically improved stocks has been increased .
Understanding population structure and genetic diversity within and between wild (W ) and farmed (F ) populations is of paramount importance to develop optimal strategies for the conservation of W populations and to achieve sustainable aquaculture production of Mediter ranean seabream and seabass. In particular, t he success of any breeding programme critically depends on the way in which the base population of breeders is built and on the control of inbreeding ( or equivalently, the effective population size, Ne) o nce selection programmes are running. Also, although l ittle is known on escapes from seabream and seabass, they pose an ecological risk of transferring diseases to wild fish and may cause undesirable genetic effects in native populations due to interbreeding .
Recently , a combined SNP array for both species has resulted from the collaboration between the EU projects MedAID and PerformFISH and is an essential tool for genetic studies for estimati ng Ne and relationships between populations (Peñaloza et al. 2021). The objectives of this study were to i ) identify the extent of W and F population stratification within species considering the geographic origin of individuals; and ii) assess the genetic status of W and F populations through the estimation of current Ne.
Material and Methods
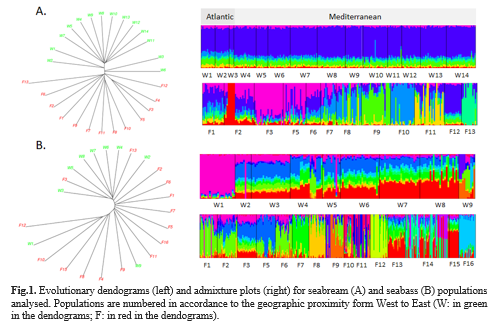
S amples were collected from W and F populations of both species from East to West Mediterranean. Three populations of seabream from the Atlantic were also sampled. For seabream, SNP genotypes were available for 462 individuals sampled from 14 W and 12 F populations . For seabass, SNP genotypes were available for 516 individuals collected from 9 W and 15 F populations. After quality filtering the total number of SNPs available for analysis was 25,319 (seabream) and 22,507 (seabass) . Population structure was assessed through (i) Principal Component Analysis (PCA) u sing the software PLINK (Purcell et al. 2007) and a R script; (ii) phylogenetic analysis using hierarchical methods with the R-package; (iii) ancestry clustering using ADMIXTURE (Alexander and Lange, 2011); and (iv) p airwise FST using Metapop2 (López-Cortegano et al. 2019) . Current estimates of Ne were obtained from linkage disequilibrium between independent SNPs as implemented in the software NeEstimator (Do et al., 2014).
Results
Results from PCA did not show a clear differentiation between W and F populations , and showed a considerably dispersion between populations. However, clustering methods (i.e. phylogenetic and admixture analysis) and FST revealed a clear differentiation between W and F populations. Despite the little differentiation among W populations, Atlantic/Mediterranean (seabream) and West/East Mediterranean ( seabream, seabass) patterns were detected (Fig. 1) . In general, Ne was large (> 1000) for W and small (< 100) for F populations of both species, with some exceptions. Farmed populations were more heterogenous and presented in many cases critical values of Ne. Average FST was lower between W (0.020 for seabream, 0.047 for seabass ) than between F (0.033 for seabream and 0.051 for seabass) populations.
Discussion
These results are in line with previous literature exploring genetic variation and population structure of W and F seabream popula tions with traditional markers, suggesting that W populations are more similar among them and present higher genetic variability and Ne than F populations (see Crosetti et al., 2014), and that there is a subtle but significant levels of population structure. These results are in accordance with results from the EU project AQUATRACE. In W populations, a certain degree of differentiation between Atlantic and Mediterranean basins has also been previously reported, as well as within Mediterranean populations. The low differentiation between W populations indicates that considerable gene flow exists , facilitating sampling decisions when base populations are established. These results also indicate that there are no specific requirements for conservation of W populations . The low Ne estimated for some F populations highlight the need of applying measures to increase this size to ensure the sustainability of the breeding programmes. Finally , since F populations are more differentiated than W , escapes from farms should be avoided. In summary, this study evidence the high potential of the genomic tool developed for establishing and monitoring supportive breeding programmes in aquaculture.
References
Alexander DH, Lange K (2011). Enhancements to the ADMIXTURE algorithm for individual ancestry estimation. BMC Bioinformatics 12:246.
Crosetti (et al., 2014 ). GIlthead sea bream (Sparus aurata ) - AquaTrace Species leaflet. D. Crosetti , AR. Rossi, S. de Innocentiis (eds).
Do C et al.. (2014). NeEstimator V2: re-implementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Resour. 14, 209-214.
López-Cortegano et al. (2019). Metapop2: Re‐implementation of software for the analysis and management of subdivided populations using gene and allelic diversity. Mol. Ecol. Resour. 19(4):1095-1100.
Peñaloza C, et al. (2021). Development and validation of a combined species SNP array for the European seabass (Dicentrarchus labrax) and gilthead seabream (Sparus aurata). Genomics. DOI:10.1101/2020.12.17.423305.
Purcell et al (2007) . PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses Am. J Hum. Genet. 81(3):559-75.