ENABLING PRODUCTION OF THE RARE CAROTENOID DIATOXANTHIN AT INDUSTRIAL LEVEL USING CRISPR/CAS9 GENE-EDITED MICROALGAE
Introduction
Recent research has indicated that the rare carotenoid diatoxanthin (Dtx) exhibits higher bioactivity than commercially available pigments, and that it has anti-inflammatory and anti-cancer properties (Konishi et al., 2008; Pistelli et al., 2021; Sansone et al., 2025, 2023, 2022). These findings highlight Dtx as a new marine compound of commercial interest. Dtx is only found in a few groups of microalgae where it is produced as protection against exposure to sudden high light intensity, and it can be viewed as a “sunscreen” pigment (Goss and Lepetit, 2015). Large-scale commercial production of this pigment from microalgae is currently not feasible. In fact, Dtx is rapidly converted to the inactive pigment diadinoxanthin (Ddx) when the cells are removed from the high light source (Goss and Lepetit, 2015; Græsholt et al., 2024), which will be the case during large-scale harvesting of algal biomass. In addition, producing commercially interesting levels of Dtx in “natural” microalgae will depend on exposing the algae culture to high intensity light for a prolonged period of time. This can be technically challenging at high cell densities and costly in countries where algae production is dependent on artificial light. Using the gene editing technique CRISPR/Cas9 we have created non-transgenic zep3Cpsrp54 double knockout (KO) mutants of the marine diatom Phaeodactylum tricornutum. The ZEP3 gene encodes zeaxanthin epoxidase responsible for back-converting high light-accumulated Dtx to Ddx in low light, whereas the CpSRP54 gene encodes the chloroplast signal recognition particle 54 kDa (CpSRP54) protein involved in insertion of subunits of photosynthetic complexes into thylakoid membranes (Græsholt et al., 2024; Nymark et al., 2021). We have previously shown that loss of ZEP3 can stabilize high light-accumulated Dtx, whereas absence of CpSRP54 causes light sensitivity (Græsholt et al., 2024; Nymark et al., 2021). The loss of both ZEP3 and CpSRP54 in P. tricornutum was hypothesised to result in a Dtx production line where both the above-described challenges would be solved. Producing a high-value product like Dtx can cause microalgae cultivation to become economically viable, opening the possibility of using the rest of the algal biomass for feed purposes.
Material and Methods
Zep3Cpsrp54 double KO mutants were generated by introducing mutations in the CpSRP54 gene in the previously published zep3 mutant lines using the CRISPR/Cas9 technology (Græsholt et al., 2024; Sharma et al., 2018). Bacterial conjugation was chosen as the method for delivery of the components necessary for CRISPR/Cas9-gene editing to take place to avoid incorporation of “foreign” DNA into the genome of the P. tricornutum cells (Sharma et al., 2018). The phenotype of the zep3Cpsrp54 double KO mutant was characterized and compared to the phenotype of zep3 and Cpsrp54 single mutants and wild type (WT) cells. The phenotypic comparisons were performed by assessing their photophysiological performances using Pulse-amplitude modulated (PAM) fluorometry, pigment concentrations by HPLC and growth rates as a daily increase in in vivo chlorophyll a fluorescence as a response to different light treatments.
Results and Discussion
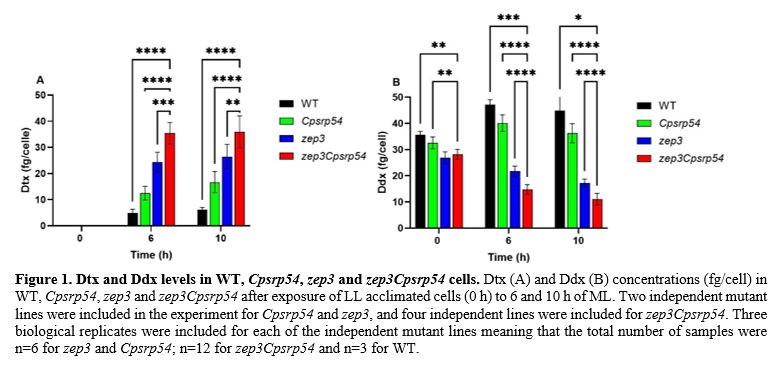
Non-transgenic zep3Cpsrp54 double KO mutants with bi-allelic mutations were successfully generated using the CRISPR/Cas9 technology. The double KO mutants were found to display the photophysiological features of both zep3 and Cpsrp54 single mutants. The double KO mutants were unable to relax their Dtx-dependent photoprotective response in low light, a feature previously observed in the zep3 single mutants (indicating that high light-accumulated Dtx was stabilized), and they were found to be light sensitive similar to the Cpsrp54 single mutants (indicating that Dtx can be produced without the use of high intensity light) (Græsholt et al., 2024; Nymark et al., 2021). To test if the zep3Cpsrp54 double mutants could function as a Dtx production line and outperform the single mutants and WT cells, we exposed low light (LL; 35 µmol photons m-2 s-1) acclimated mutants and WT to medium light (ML; 200 µmol photons m-2 s-1) for 6 and 10 h. Pigment analysis performed on material harvested from LL acclimated cultures and 6 h and 10 h ML treated cultures showed that ML was enough to trigger the double KO mutants to produce high levels of Dtx (Figure 1A). The highest concentrations of Dtx were found in the double KOs, followed, in decreasing order, by zep3, Cpsrp54 and WT (Figure 1A). The opposite trend was observed for the Ddx concentration (Figure 1B). The Dtx production in the double KOs clearly outperformed the production in the zep3 lines showing Dtx concentrations that were 46% and 36% higher than in zep3 lines after 6 h and 10 h of ML treatment, respectively. The Dtx concentration in the double KO mutants after 6 h in ML reached similar levels as previously reported for zep3 single mutants exposed to high intensity light (500 µmol photons m-2 s-1) (Græsholt, 2024). Even though the concentration of Dtx differed significantly between mutant lines, they were highly similar for the same mutant line at both 6 h and 10 h (Figure 1A). An additional experiment using shorter ML exposure times (0.5 h and 2 h) showed that prolonged exposure is necessary to reach maximum Dtx levels. Growth rates of the double KO mutants were at WT levels up to 150 µmol photons m-2 s-1 supporting the idea that these mutants could function as a production line of Dtx. Future project work will include large-scale cultivation of zep3Cpsrp54 double KO mutants together with our industry partners to prove that commercial production of Dtx is a possibility.
References
Goss, R., Lepetit, B., 2015. Biodiversity of NPQ. J Plant Physiol 172, 13–32.
Græsholt, C., Brembu, T., Volpe, C., Bartosova, Z., Serif, M., Winge, P., Nymark, M., 2024. Zeaxanthin epoxidase 3 knockout mutants of the model diatom Phaeodactylum tricornutum enable commercial production of the bioactive carotenoid diatoxanthin. Marine Drugs 22, 185.
Konishi, I., Hosokawa, M., Sashima, T., Maoka, T., Miyashita, K., 2008. Suppressive effects of alloxanthin and diatoxanthin from Halocynthia roretzi on LPS-induced expression of pro-inflammatory genes in RAW264.7 cells. J. Oleo Sci. 57, 181–189.
Nymark, M., Hafskjold, M.C.G., Volpe, C., Fonseca, D.D., Sharma, A., Tsirvouli, E., Serif, M., Winge, P., Finazzi, G., Bones, A.M., 2021. Functional studies of CpSRP54 in diatoms show that the mechanism of thylakoid protein insertion differs from that in plants and green algae. Plant J 106, 113–132.
Pistelli, L., Sansone, C., Smerilli, A., Festa, M., Noonan, D.M., Albini, A., Brunet, C., 2021. MMP-9 and IL-1β as targets for diatoxanthin and related microalgal pigments: Potential chemopreventive and photoprotective agents. Mar. Drugs 19, 354.
Sansone, C., Pistelli, L., Brunet, C., 2025. The marine xanthophyll diatoxanthin as ferroptosis inducer in MDAMB231 breast cancer cells. Sci Rep 15, 8146.
Sansone, C., Pistelli, L., Calabrone, L., Del Mondo, A., Fontana, A., Festa, M., Noonan, D.M., Albini, A., Brunet, C., 2023. The carotenoid diatoxanthin modulates inflammatory and angiogenesis pathways In vitro in prostate cancer cells. Antioxidants 12, 359.
Sansone, C., Pistelli, L., Del Mondo, A., Calabrone, L., Fontana, A., Noonan, D.M., Albini, A., Brunet, C., 2022. The microalgal diatoxanthin inflects the cytokine storm in SARS-CoV-2 stimulated ACE2 overexpressing lung cells. Antioxidants 11, 1515.
Sharma, A.K., Nymark, M., Sparstad, T., Bones, A.M., Winge, P., 2018. Transgene-free genome editing in marine algae by bacterial conjugation – comparison with biolistic CRISPR/Cas9 transformation. Sci Rep 8, 14401.