CENTRAL INFLAMMATORY RESPONSE IN RAINBOW TROUT AND ITS IMPLICATIONS IN APPETITE REGULATION
Introduction
Increased dietary fat has been used in aquafeeds to enhance fish growth while reducing costs, as fats are energy rich and cheap (Naiel et al., 2023). However, in mammals, high-fat diets (HFD) seem to trigger a generalized inflammatory response that alters fee d intake regulation mechanisms (Le Thuc et al. , 2017), leading to hyperphagia (Lee et al. , 2020), linking inflammation and feed intake processes . In mammals, dietary fat triggers a rapid hypothalamic inflammatory response 24 h after the first feeding of HFD and reaches a peak around the third day, descending for 2-3 weeks, and rising again becoming chronic and systemic (Thaler et al. , 2013; Jais y Brüning, 2017, Lee et al. , 2020). Mechanistically, two major inflammatory pathways are activated: those mediated by JNK and NF-κB. As a result, proinflammatory cytokines are produced (Le Thuc et al. , 2017). Additionally, oxidative stress seems to be impaired because of toxic lipidic species accumulation (Cavaliere et al. ). Saturated fatty acids seem to be the trigger of this complex response (Chataigner et al., 2021). In fish, little is known about the relationship of feeding with inflammatory response associated to dietary fat. The inflammatory response has been mostly studied at peripheral levels and central effects have been demonstrated in fish (Dai et al., 2018 ; Ghaddar et al., 2020). The purpose of this study was to determine if the central inflammatory response to dietary fat observed in mammals is conserved in a teleost fish and if it is also related with feed intake.
Materials and methods
60 rainbow trout (Oncorhynchus mykiss) were randomly distributed into 4 equal tanks and fed a commercial diet (Biomar, Spain) during the acclimatation period. Then, two tanks were fed the control diet (CD), whereas the other two were fed the high-fat diet (HFD), with a 24 and 32 (% DM) of fat content, respectively. Fish were fed until visual satiety once a day and feed intake levels were recorded. 3 and 17 days after eating the experimental diets (Sparos, Portugal), fish were anaesthetized and euthanized to collect blood, hypothalamus and telencephalon, that were immediately frozen in dry ice and stored at -80 ºC until further analysis. Glucose, lactate, fatty acids and triglycerides concentrations were measured in plasma. The relative amount of mRNA from genes related with inflammatory response, oxidative stress and feed intake regulation in hypothalamus and telencephalon was quantified by RT-PCR. Statistical analysis was performed to all data using R and RStudio software. ANOVA, Tukey tests and non-parametric Kruskall-Wallis tests were done as needed.
Results
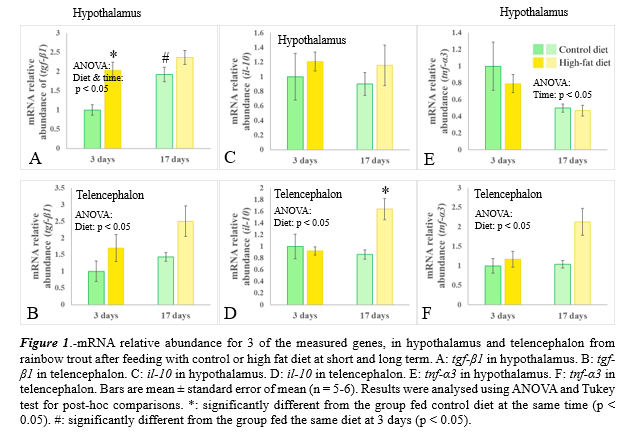
HFD led to a significantly higher plasmatic fatty acid concentration. An increasing trend was also observed in plasmatic triglycerides concentration. Antiinflammatory cytokines (tgf-β1 and il-10) mRNA relative abundance showed a general tendency to increase over time in both (hypothalamus and telencephalon) (Fig. 1A-D) . Regarding proinflammatory cytokines, tnf-α mRNA relative abundance increased at long term in HFD group in telencephalon, whereas in hypothalamus, there is a decreasing trend in short term (Fig. 1E-F) . Genes related to inflammatory pathways (ikkα and ikkβ ) showed an increasing tendency at 17 days in both tissues in HFD group. All three oxidative stress related genes mRNA relative abundance showed an increasing trend in both tissues at long term (data not shown). As for feed intake regulation, hypothalamic anorexigenic neuropeptides (pomca1, cartpt ) mRNA relative abundance tended to increase at long term in HFD. Orexigenic neuropeptides (npy and agrp) mRNA abundance showed an increasing tr end in those animals fed the HFD at short term in the hypothalamus (data not shown).
Discussion and conclusions
Inflammatory response seemed to be stronger and more consistent at telencephalon level attending to mRNA relative abundance variations. The increasing tendency observed in genes involved in inflammatory pathways along with the changes observed in pro- and anti-inflammatory genes support the hypothesis of a conserved inflammatory response to HFD at central level in teleost fish as observed previously in mammals (Souza et al. , 2016; Cavaliere et al. , 2018). Additionally, the pattern of variation displayed suggests that the sampling times, selected in accordance with the pattern observed in mammals, were not the optimal to catch the peak of inflammation, that could be due to immune responses being slower in fish (Watts et al., 2001). On the other hand, a long-term orexigenic response was observed.
In summary, results suggest the existence of an inflammatory response against high fat diets that develops slower than in mammals. Moreover, this response seems to be related with homeostatic regulation of feeding, and could also be related to hedonic control, since telencephalon is the main brain area affected. Further studies are needed to address both the implication of hedonic control and the temporal pattern of inflammatory response.
Bibliography
Cavaliere, G.; Viggiano, E.; Trinchese, G.; De Filippo, C.; et al (2018). Frontiers in physiology, 9, 818.
Chataigner, M.; Martin, M.; Lucas, C.; Pallet, V. ; et al (2021). Nutrients , 13, 824.
Dai, Y. J.; Jiang, G. Z.; Yuan, X. Y.; & Liu, W. B. (2018). British Journal of Nutrition , 120, 1422-1431.
Ghaddar, B.; Veeren, B.; Rondeau, P.; Bringart, M. ; et al (2020). Scientific Reports , 10, 14496.
Jais, A.; & Brüning, J. C. (2017). The Journal of clinical investigation , 127, 24-32.
Le Thuc, O.; Stobbe, K.; Cansell, C.; Nahon, J. L.; et al (2017). Frontiers in Endocrinology, 8, 197.
Lee, C. H.; Suk, K. ; Yu, R.; & Kim, M. S. (2020). Molecules and cells , 43, 431-437.
Naiel, M. A.; Negm, S. S.; Ghazanfar, S.; Shukry, M.; et al . (2023). Journal of Animal Physiology and Animal Nutrition , 107, 948-969.
Souza, G. F.; Solon, C.; Nascimento, L. F.; De-Lima-Junior, J. C.; et al (2016). Scientific reports , 6, 29290.
Thaler, J. P.; Guyenet, S. J.; Dorfman, M. D.; Wisse, B. E.; & Schwartz, M. W. (2013). Diabetes , 62, 2629-2634.
Watts, M.; Munday, B. L.; & Burke, C. M. (2001). Australian Veterinary Journal, 79, 570-574.