EFFECTS OF PRODUCTION TEMPERATURE, WATER CURRENT VELOCITY, AND THEIR INTERACTION ON FARMED ATLANTIC SALMON Salmo salar STRESS RESPONSES AND WELFARE
Introduction
Farmed Atlantic salmon (S. salar) mortalities have increased in recent years, occurring during both the freshwater (FW) and seawater (SW) production phases. Meanwhile, suboptimal rearing and transfer strategies between FW and/to SW can greatly influence fish resilience. For instance, seasonal mismatches in the thermal environment during transfer can have pronounce effects on fish welfare and challenge abilities to adjust to the novel SW environment. At the same time, high water velocities are increasingly favoured in aquaculture settings due to reported benefits on fish health, including improved stress coping and cardiovascular fitness. However, the interactions between such training procedures and temperature and their effects on stress functions critical for farmed salmon welfare is still poorly known. Therefore, this study aimed to evaluate how temperature, water current velocity and their interaction influences farmed salmon welfare by monitoring stress responses and key neural markers of brain plasticity, with special reference to the sensitive FW-SW production phase.
Materials and methods
Atlantic salmon parr (initial body weight ̴ 40g) were allocated in duplicates between two water velocities (body length [BL] / second [s]): 1) H (1.2 BL/s) and 2) L (0.6 BL/s). The fish were maintained under these conditions at 10°C in FW RAS for 8-weeks. During this period fish were smoltified according to standard industry practice, followed by SW transfer of smolts in the same respective water velocities. To investigate the effect of abrupt temperature changes that often results from transfer from land-based sites to sea-cages, we investigated three distinct temperature regimes (1) 8°C [8-H & 8-L], 2) 10°C [10-H & 10-L] and 3) 14°C [14-H & 14-L]). The fish remained under these conditions for an additional 8-weeks before final sampling. At the end of both FW and SW phases, 20 fish from each treatment (N=5 per tank) were sampled for blood and brain tissue. In each treatment, 10 fish were immediately euthanized (pre-stress) and the other 10 were subjected to an acute challenge test (ACT), by high density, to monitor stress response capacities (ACT; 15 min. at 300 kg. m-3, followed by 45 min. recovery). Plasma levels of cortisol were measured using ELISA, and ions (e.g., sodium, chloride) and energy metabolites (e.g., glucose, lactic acid) analysed using Horriba Pentra 400. These were compared to telencephalic expression changes in genes involved in the stress response (mineralocorticoid receptor; mr, glucocorticoid receptors; gr1 and gr2) and neural plasticity (e.g., neurogenic differentiation factor; neurod, e.g., immediate early gene [IEG]; c-fos). Data was subjected to one- and two-way ANOVAs (p < 0.05).
Results & Discussion
Effect on growth performance: Fish final average body weight was ̴ 415 g. In FW, the specific growth rate (SGR) was unaffected by water velocity. In SW, SGR increased towards higher temperature regimes (8°C: ̴ 1.61, 10°C: ̴ 1.68, 14°C: ̴ 1.86). Only at 14°C was the SGR further improved using high-water velocity (1.2 b/s).
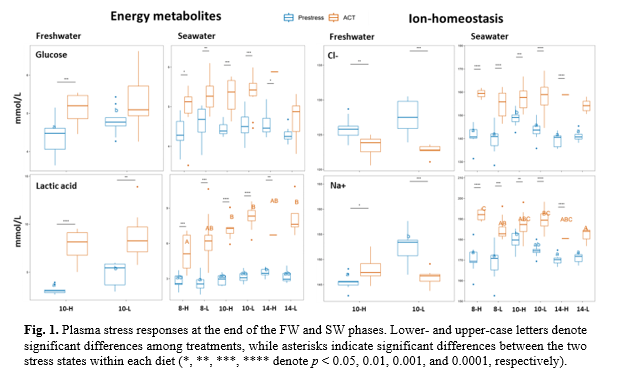
Plasma ions and metabolites: In FW, plasma glucose, lactic acid, and Na+ levels were lower in response to the high-water velocity (Fig. 1). This may indicate larger energetic demands and disturbed ion-homeostasis, effects consistent with osmo-respiratory compromise. In SW, the pre-stress 10-H group at 10°C showed a general elevation in plasma ions suggesting increased osmoregulatory challenges (Fig.1). Conversely, lactic acid levels were highest in the 14°C group following the ACT, while the high-water velocity seemed to elevate lactic acid build-up at pre-stress status in the 14-H group (Fig. 1). This build-up in lactic acid may be a consequence of thermal increases in metabolic rate alongside higher energetic demands to sustain swimming activities that elevate anaerobic forms of metabolism.
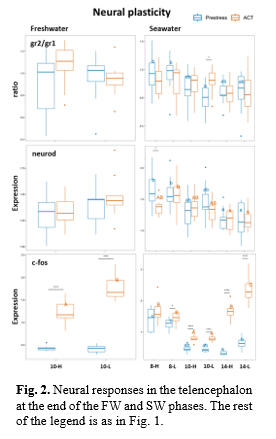
Neural functions: A gradual suppression in corticosteroid receptors (mr, gr2) and neural plasticity (neurod) was observed with increasing temperature, consistent with our prior findings (Fig. 2) (Tang et al., 2022). However, the effect of water current velocity was less apparent. In FW, the response of c-fos to the ACT appeared lower in the fish that had experienced the highest velocity strength, which could suggest reduced stress coping, while, in SW, this difference was only observed at the highest temperature that agrees with the more stressful effects seen in plasma lactic acid levels in the 14-H group.
Conclusion: The effect of water velocity in SW was less apparent compared to that of temperature. Since growth was maintained well throughout all treatments, this suggests that farmed salmon still handled the tested temperature-water velocity treatments well without marked effects on welfare status.
References
Tang et al., 2022. https://doi.org/10.3389/fmars.2022.926136
Acknowledgments
Project (328724) funded by Norwegian Research Council