TRANSCRIPTOMIC PROFILING OF FOLLICULAR CELLS ACROSS THE ANNUAL REPRODUCTIVE CYCLE IN EUROPEAN SEA BASS Dicentrarchus labrax
Introduction
Reproductive control and prevention of precocious puberty are key for sustainable aquaculture. The European sea bass (D. labrax) is economically important in the Mediterranean, yet the molecular regulation of its oogenesis remains incompletely understood. Oocyte development involves systemic hormonal cues (FSH, LH, sex steroids) (1) and local paracrine signals within the ovarian follicle, between the oocyte and surrounding granulosa cells (2). Follicular cells support oocyte growth and maturation, and coordinate folliculogenesis through specific signaling pathways (3). This study aimed to investigate seasonal changes in gene expression in follicular cells over a full reproductive cycle to identify the molecular mechanisms driving follicular development (4)
Materials and methods
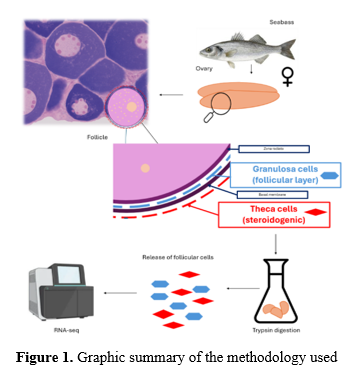
Follicular cells were isolated throughout the annual cycle from ovaries of adult females kept under natural photoperiod and temperature at the Instituto de Acuicultura Torre de la Sal (IATS), following Crespo et al. (5). Ovaries were dissected, washed, and dissociated in M199:H₂O medium with antibiotics. Enzymatic digestion was performed with trypsin (37 °C, 30 min), followed by filtration (100 μm), centrifugation, and resuspension in M199 Hanks medium. Follicular cells were counted using Trypan blue in a Neubauer chamber. RNA was extracted with the Maxwell® LEV simplyRNA Tissue kit, and quality was assessed with Nanodrop® and Bioanalyzer. Libraries were prepared and sequenced (Illumina NovaSeq X) by BMKgene.
Results
A total of 67 genes involved in steroid hormone biosynthesis were identified. The highest number of differentially expressed steroidogenic genes occurred in spring/summer versus winter, with 13 and 15 DEGs (mostly upregulated), respectively. These data highlight dynamic seasonal regulation of gene expression in follicular cells, particularly in relation to steroidogenesis and follicle development.
Transcriptomic profiles were compared using RNA-seq of follicular cells isolated from female European sea bass ovaries collected during the four seasons (summer, autumn, winter, and spring). Ovarian staging was performed by histological analysis of each ovary and classified as previtelogenic, vitelogenic, maturing/ovulating and atresic. Major transcriptional changes were observed between late winter (end of the spawning period) and summer (post-spawning), coinciding with follicular atresia and initiation of new follicle formation, so many of these differentially expressed genes are probably related to folliculogenesis. A second transcriptional shift was noted at the onset of vitelogenesis respect to the previtelogenic stage, while few changes occur during the secondary growth from initial vitellogenesis to postvitellogenesis / maturation. A total of 67 genes involved in steroid hormone biosynthesis were identified. The highest number of differentially expressed steroidogenic genes occurred in spring/summer (previtelogenesis) versus winter (spawning), with 13 and 15 differentially expressed steroidogenic genes (mostly upregulated), respectively. These data highlight dynamic seasonal regulation of gene expression in follicular cells, particularly in relation to steroidogenesis and follicle development.
Discussion and Conclusion
Transcriptomic analysis of follicular cells revealed two main peaks of gene expression. An increase in gene expression activity after the spawning period (late spring / summer), likely related to tissue remodeling and folliculogenesis (7). Another peak in gene expression upregulation could be observed in winter, at the beginning of the spawning phase. This last pattern aligns with previous findings on key reproductive genes (fshr, lhr, star, cyp19a1a) during winter in European sea bass ovaries (6). Among steroidogenic genes, cyp11a1.2 and hsd3b1 were consistently upregulated across seasons. In conclusion, these results demonstrate that follicular cell transcriptional activity in D. labrax is seasonally regulated, contributing to the understanding of ovarian development and informing improved reproductive strategies in aquaculture.
Funding
Funded by MCIN/AEI/10.13039/501100011033/ and by ERDF a way of making Europe (PID2021-122929OB-C32). A.M. and A.E. are supported by PhD contracts from GV (GRISOLIAP/2020/129, CIGRIS/2022/002).
References
- Ge, W. (2005). Fish Physiol. Biochem. 31, 209–214
- Lubzens, E. et al. (2010). Gen. Comp. Endocrinol. 165(3), 367-389
- Clelland, E. et al. (2009). Mol. Cell. Endocrinol. 312(1-2), 42-52.
- Zayed Y, et al. (2019) . Front. Endocrinol. 10:518
- Crespo B, et al. (2013). Cytotechnology. 65:273–286
- Rocha, A. et al. (2009). Gen. Comp. Endocrinol. 162(3), 265–275.
- García-López, A. et al. (2011). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 160(3), 374–380.