TURN-UP THE SHELL: ADVANCES ON THE PATELLID LIMPETS' AQUACULTURE
Introduction
The true limpets (Patellogastropoda ) are highly specialized marine gastropods commonly found colonizing rocky seashores . The limpets have been a fishery resource since the Neolithic period . Currently several limpet species are threatened by the overexploitation, which is leading to smaller and male biased populations , which individuals have less reproductive capacity and higher inbreeding ratio. Consequently, several limpets populations are endangered being taken protective measures focused on harvesting restrictions
(Sousa et al., 2020)
. As alternative measure for the exploitation of the natural stocks, the limpets’ a quaculture could be a sustainable effort able to develop different programs , either for human consumption or for the reproduction and reintroduction of different limpet species.
Limpets’ aquaculture is a recent research field which efforts were mainly focused on the larvae production with variable success, while juvenile production has been mostly unsuccessful thus far. Recently, this state of the art changed thanks to advances done in Madeira Island and focused on the native species Patella aspera and P. candei , it is now possible to obtain juveniles from an already settled larvae production .
Material and Methods
The adults were captured during the breeding period (mid autumn to mid spring in both species ) and kept using an open system with high aeration, at 20 ± 1 °C and 36 ± 1 psu . The adults showed higher survival when no lesions occurred in the soft body during capture and management in captivity. Gametes were obtained dissecting a dult specimens to expose the gonads . The sperm was gathered using a Pasteur’s pipette and diluted in 100 ml filtered seawater. The sperm of 4 males was pooled and kept in the fridge until required. The oocytes were released breaking a female gonad placed inside a crystal beaker with filtered seawater using a Pasteur’s pipette. Then, the oocytes were carefully washed using nylon meshes (200 and 55 µm). The oocytes of 4-5 females were pooled and matured artificially using NaOH alkalinized seawater baths during 3h (pH 9.0 in P. aspera and 9.5 in P. candei) . The oocytes should be carefully washed after maturation since longer baths could damage the oocytes. The matured oocytes were fertilized at 100 oocytes ml-1 (or less) and 105 sperm cells ml-1 f or a better larval production. Fecundation was realized in 500 ml crystal beakers. The sperm was kept with the oocytes during the incubation time, which lasted 24h.
Larvae were observed 24h post-fecundation as trocophora , 2/3 of the water column was siphoned using a 55 µm filter to avoid collecting debris, then the larvae were re-distributed in new beakers with clean filtered seawater at 10-20 larvae ml-1, and cultured during additional 48h until pediveliger stage was reached. Then, the larvae were siphoned again using a 55 µm filter and re-distributed in culture cell plates at 5-10 larvae ml-1 for experimental purposes. Several substrates plus control were tested: two diatom species (Halamphora and Navicula ), haptophyte Pavlova , and limpet shells covered by different communities of crustose coralline algae (CCA) as natural substrate. The timing for settlement was based on the observation of post-larvae, while success for settlement was based on the average ratio of juveniles in each treatment. The incubation, larval culture, and settlement assays were realized at 17 ± 1 °C.
Results and Discussion
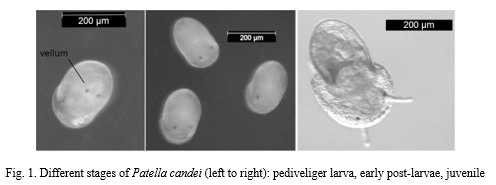
Post-larvae were identified by the shedding of the velum and increased eye distance, while juveniles showed juvenile shell and active grazing behaviour . Earliest post-larvae occurred on CCA treatments in both species, while settlement success varied in both species: P. aspera juveniles were observed on CCA treatments (with rare exceptions) , while P. candei juveniles were more frequent on Navicula biofilms and CCA treatments . The type of CCA community and their abundance influenced the settlement success, being P. aspera apparently more restrictive than P. candei on their settlement requirements. The larval development was completed in absence of food suggesting lecithotrophy and facilitating the larvae management. On the contrary, juvenile were active feeders. G razing was concomitant with movement, which suggest that limpet juveniles are opportunistic consumers that needs continuous feeding . The development of a juvenile aquafeed is a mandatory step for further studies on the limpets’ aquaculture. In this sense, t he observation of P. candei juveniles grazing upon Navicula biofilms marks a starting point for the culture of the limpet juveniles.
Bibliography
Castejón, D., Cañizares, J.M., Nogueira, N., Andrade, C.A.P., 2021. Artificial maturation and larval production of the limpet Patella aspera Röding, 1798 (Patellogastropoda, Mollusca): Enhancing fertilization success of oocytes using NaOH-alkalinized seawater. Aquac. Res. 52, 1904– 1914.
Faria, J., Pita, A., Martins, G.M., Ribeiro, P.A., Hawkins, S.J., Presa, P., Neto, A.I., 2018. Inbreeding in the exploited limpet Patella aspera across the Macaronesia archipelagos (NE Atlantic): Implications for conservation. Fisheries Research. 198, 180-188.
Sousa, R., Pinto, A., Vasconcelos, J., Riera, R., 2020. Does harvesting affect the relative growth in Patella aspera Röding, 1798? The European Zoological Journal. 87, 395-401.