DIETARY METHYLMERCURY AND SELENIUM INTERACTIONS IN RAINBOW TROUT JUVENILES Oncorhynchus mykiss
Introduction
Mercury (Hg), especially the organic, methylated form (MeHg), is a toxic heavy metal that can accumulate in large amounts in fish through time in various organ s, like liver and brain. Brain, with neurons, is the pri ncipal target of MeHg toxicity (Ni et al. 2012 ). MeHg induces its toxicity through induction of oxidative stress and inflammation
. Selenium (Se), a n essential micronutrient mainly involved in antioxidant protection , has been shown to reduce MeHg toxicity. However, Se and MeHg interaction s are far to be totally understood. As relatively high levels of these two elements can be found in marine ingredients, a better understanding of their interactions is important to improve aquafeed formulations .
Material and Methods
Six plant protein-rich diets were formulated to contain two different MeHg levels (0 and 2.5 mg Hg/kg in HgC and HgH diets ) combined with 3 different Se levels and forms (0.3 mg Se/kg in SeC diets and 2.6 mg Se/kg supplied either as sodium selenite in SeI diets or as L-selenomethionine from Orffa in SeO diets). Each diet was distributed to 3 replicate tanks of 50 fish (initial mean body weight: 26 ±1 g) to apparent satiation over a 3-week feeding trial at a water temperature of 17.5°C. Fish g rowth performance (n=3 tanks/diet) was analysed as well as total Hg and Se contents (n=3 individuals/diet) and relative gene expression levels (RT-PCR, n=9 individuals/diet ) in liver and brain.
Results
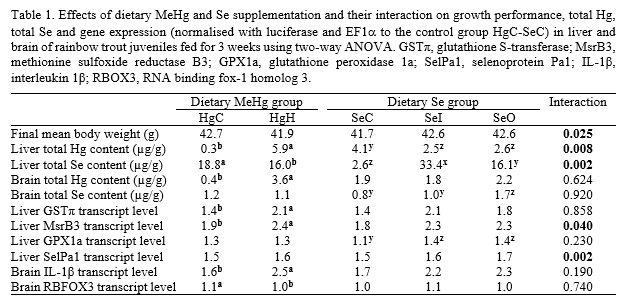
Reduced growth performance of fish fed MeHg was recorded only within the non-Se supplemented SeC-group (Table 1). Feeding MeHg significantly increased total Hg concentrations in both liver and brain but to a significant smaller extent in liver when fish were supplemented with Se. Along with the hepatic increase of total Hg, dietary MeHg supplementation decreased total Se content in liver. Dietary inorganic Se supplementation increased total Se concentrations in liver to a larger extent than dietary organic Se supplementation but inorganic Se intake did not increase significantly Se content in brain contrary to organic Se intake.
In liver, MeHg intake increased transcript levels of enzymes involved in cellular stress response and antioxidant defences such as GSTπ and MsrB3 (Table 1) . Both dietary organic and inorganic Se supplementation increased hepatic transcript levels of the selenoprotein GPX1a and also SelPa1 but only in HgC groups. In brain, dietary MeHg supplementation decreased transcript levels of RBFOX3, a neural marker, and up-regulated expression of IL-1β, a pro-inflammatory cytokine.
Discussion and conclusion
The growth inhibi tion by MeHg after only 21 days of feeding could be due to the prooxidant and proinflammatory properties highlighted by gene expression analysis in brain and liver. Fighting against MeHg-induced oxidative stress and inflammation consumes energy that is thus less available for growth (Morcillo et al. 2017). The mitigation by dietary Se supplementation that promoted antioxidant genes lends further support to this hypothesis.
Indeed, at the transcript level we found that MeHg induced inflammation in brain and antioxidant defences in liver to fight against oxidative stress . The antioxidant support by d ietary Se supplementation in plant-based diets at the transcript level, in agreement with previous studies (Fontagné-Dicharry et al., 2020), could provide better protection against xenobiotics like MeHg in rainbow trout juveniles , even if the controlled genes by dietary MeHg and Se are not systematically similar and deserved further investigation for a better understanding.
The present results provide evidence of interactions between dietary MeHg and Se on tissue Hg and Se concentrations, especially in liver. The difference of Hg and Se storage between the two tissues suggest post-absorptive interacting effects between Hg and Se maybe at the depuration level due to their strong affinity for each other as suggested by Amlund et al. (2015) in zebrafish but do not exclude interactive effects at the digestive or absorption level that deserve further investigation. As muscle, the edible part, has been shown to readily accumulate MeHg (Berntssen et al. 2004), this tissue should be included in the study to ascertain this finding with different dietary Se levels and forms.
Our data showed that the dietary level of 2 mg Hg/kg diet supplied as organic mercury in duced adverse effects as early as 21 days of feeding, especially in Se-deficient diets , in agreement with the threshold levels of dietary MeHg set as 5 mg Hg/kg by Berntssen et al. (2004) for salmonid diets. In brain, our RBFOX3 results confirm the high sensitivity of neurons to MeHg toxicity (Ni et al. 2014). Longer exposure, for example up to 6 months, should be assessed in rainbow trout juveniles for a better understanding on the adverse effects of MeHg and the interacting effects with dietary Se.
Acknowledgements
This project was supported by the French National Research Agency (ANR-18-CE34-0004), the Region Nouvelle-Aquitaine (BENESEL) and I-site E2S UPPA (contract 2020-32) and has received funding from the European Union’s Horizon 2020 research and innovation programme under the Marie Sklodowska- Curie grant agreement No. 101007962.
References
Amlund , H., Lundebye , A.-K., Boyle, D., Ellingsen , S., 2015. Dietary selenomethionine influences the accumulation and depuration of dietary methylmercury in zebrafish ( Danio rerio ). Aquat . Toxicol . 158: 211-217.
Berntssen , M.H.G., Hylland , K., Julshamn , K., Lundebye , A.-K., Waagbø , R., 2004. Maximum limits of organic and inorganic mercury in fish feed. Aquac . Nutr . 10: 83-97.
Fontagné-Dicharry , S., Véron , V., Larroquet, L., Godin, S., Wischhusen, P., Aguirre, P., Terrier, F., Richard, N., Bueno, M., Bouyssière , B., Antony Jesu Prabhu, P., Tacon, P., Kaushik, S.J., 2020. Effect of selenium sources in plant-based diets on antioxidant status and oxidative stress-related parameters in rainbow trout juveniles under chronic stress exposure. Aquaculture 529: 735684.
Morcillo, P., Esteban, M.A., Cuesta, A., 2017. Mercury and its toxic effects on fish. AIMS Environ. Sci. 4: 386-402.
Ni, M., Li, X., Rocha, J.B.T., Farina, M., Aschner , M., 2012. Glia and methylmercury neurotoxicity. J. Toxicol . Environ. Health A 75: 1091-1101.