BIOFORTIFIED FISH PRODUCTS FOR CONSUMERS WITH NATRUAL IODINE, SELENIUM AND FATTY ACIDS: IMPACTS ON FISH PHYSIOLOGY
Introduction
Tailor-made fortified seafood products can contribute to overcome one third of global population nutritional deficiencies, particularly on iodine, selenium and iron (1). Therefore, eco-innovative biofortified diets in iodine (macroalgae), selenium (yeast), EPA and DHA (microalgae), were enhanced with levels of these essential nutrients within the legal limits for consumers (2) and tested with gilthead seabream (Sparus aurata). These nutrients are known to interfere in vertebrates important metabolic and physiological functions, thus affecting both humans and fish. Iodine is an essential mineral for thyroid function (metabolism), selenium is involved in different and important enzymatic reactions of different pathways (deiodinases, oxidative stress, digestive enzymes, etc) and EPA and DHA are essential for membranes formation, lipid metabolism, synthesis of important bioactive molecules, among others. Therefore, biofortification of farmed fish products must assure that levels used in diets do not interfere with fish physiological functions, to avoid compromising fish health and welfare. The aim of this study was to assess the effect of the biofortified diets on different aspects of seabream physiology namely on digestive physiology, blood analysis and liver condition.
Material and Methods
Trial was carried out at IPMA’s aquaculture research station in Olhão, for 3 months, using gilthead seabream (2). Fish were fed four experimental diets manufactured by SPAROS, one based on commercial formulation (CTRL), and three enriched diets (B1, B2 and B3) supplemented with different blends of iodine-rich macroalgae (0.40% in B1 and B2; 0.80% in B3) and selenized yeast (0.015% in B1 and B2; 0.035% in B3). At the end of the trial six fish were anesthetized, blood collected from caudal vein, and afterwards sacrificed to collect intestine and liver samples for analysis. Histological sections of intestine and liver stained with H-E were made to analyse tissue structure and integrity. Aminopeptidase and alkaline phosphatase activities were determined using specific substrates. Liver fatty acid profile was determined by GC-FID. EPOC multiparameter blood reader (Siemens Healthcare) was used for blood analysis (haematocrit, haemoglobin, pH, pCO2, pO2, K+, glucose and lactate). Plasma was used for total lipids, triglycerides, and phospholipids determination using clinical diagnostic kits from SPINREACT (Spain), whereas enzyme immunoassay kits from IBL (Germany), were used for quantitative determination of triidothyronine and free triiodthyronine (fT3).
Results
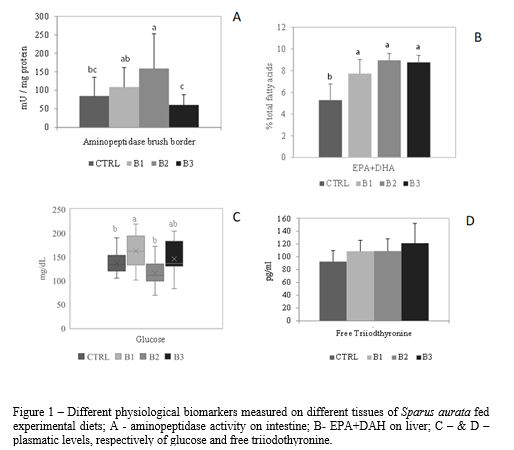
The histological structure of gilthead seabream intestine and the number of mucous cells scattered on the villi epithelium (P>0.05) were similar among the different dietary treatments. The activity of digestive enzymes was differently affected by the diet, since fish fed B2 diet exhibited higher aminopeptidase activity (P<0.05), whereas alkaline phosphatase was higher for fish fed CTRL and B3 diets. Liver histological structure of fish was similar regardless the diet. Liver fatty acid profile exhibited few differences among treatments, where higher levels of n-3 fatty acids reflected the biofortification process.
Blood analysis of gilthead seabream indicated that parameters varied within similar intervals of values. Still, pH, oxygen and glucose were higher on fish fed B1 diet (P<0.05). Similar values were observer for the analysed plasmatic parameters (lipid metabolism and thyroid hormones) among treatments, although FT3 level tended to increase in the biofortified diets.
Discussion
Physiological biomarkers values observed for gilthead seabream fed biofortified diets were largely similar when compared to control diet. Biofortified diets induced different physiological responses by gilthead seabream for the different parameters studied. The different composition of biofortified diets, manufactured with different ingredients and doses, might have triggered biological pathways differently, resulting in different physiological responses by fish. Integrating some of the physiological biomarkers (alkaline phosphatase activity and FT3 levels) with zootechnical parameters (2), suggests that B3 diet enhanced fish metabolism but with low energy efficiency, due to the higher FCR and lower weight attained at the end of the trial.
Overall the biofortification of fish farmed products for consumers is possible without compromising fish health and welfare.
Acknowledgments
The authors acknowledge funding from European Union’ s Horizon 2020 research and innovation programme under Grant Agreement no. 773400 (SEAFOODTOMORROW) and MAR2020 program through DIVERSIAQUA II project (MAR-02.01.01-FEAMP-0175).
References
1. Pinkaew and Karrila, 2015. Key properties of iodine-, iron- and zinc- fortified fish cracker: effects of ambient shelf storage on iodine retention and quality indicators. doi:10.1111/ijfs.12871
2. Barbosa V. et al., 2020. Enriched feeds with iodine and selenium from natural and sustainable sources to modulate farmed gilthead seabream (Sparus aurata) and common carp (Cyprinus carpio) fillets elemental nutritional value. doi.org/10.1016/j.fct.2020.111330