DEVELOPMENT USING BIOLUMINESCENCE IMAGING OF A RECOMBINANT ANGUILLID HERPESVIRUS 1 VACCINE CANDIDATE ASSOCIATED WITH NORMAL REPLICATION IN VITRO BUT ABORTIVE INFECTION IN VIVO
Introduction
Anguillid herpesvirus 1 (AngHV-1) (recently renamed Cyvirus anguillidallo 1) is the etiologic agent of a lethal disease that affects several eel species. It is thought to be one of the main infectious agents causing a population decline in wild eels and economic loss within the eel aquaculture sector. Since the first report of AngHV-1 in the 1980s, it has been detected globally, affecting different anguillid eel species from temperate to tropical countries. To date, there is no vaccine against AngHV-1 available on the market. Current measures to prevent AngHV-1 disease within aquaculture settings mainly rely, when possible, on shifting water temperatures outside of the range (below 22 °C) associated with efficient lytic replication. However, it promotes latent infection thereby increasing the risk of transmission when latently infected eels are cohabiting with naïve eels under stressing conditions. Recently, we developed an attenuated recombinant vaccine against CyHV-3 using prokaryotic recombination technologies and in vivo bioluminescent imaging system (IVIS). The development of this vaccine relied on the deletion of CyHV-3 ORF57. Orthologues of CyHV-3 ORF57 exist in CyHV-1 and CyHV-2 (ORF57) and in AngHV-1 (ORF 35). Therefore, this study demonstrates the effect of ORF35 deletion on the ability of AngHV-1 to replicate in cell culture and on its virulence after inoculation in European eels (yellow eel stage) by different routes.
Materials and methods
Eel kidney (EK-1) cells were cultured. The AngHV-1 isolate (UK strain) was used as a parental strain to produce recombinant AngHV-1 UK Luc strain, UK ORF35 Del and UK Luc ORF35 Del by homologous directed recombination (HDR). Luc strain encodes a LucGFP (luciferase and copGFP) cassette inserted in the intergenic region ORF32–ORF33 of AngHV-1 genome. ORF35-deleted recombinants were produced by replacing ORF35 with a sequence encoding mCherry. All the recombinants in this study were confirmed by SacI restriction fragment length polymorphism (RFLP) and transcriptional analyses. Indirect immunofluorescence staining, viral growth and plaque-area assay for recombinant AngHV-1 strains were investigated. These recombinants were inoculated in eels by intraperitoneal injection or intradermic infection or immersion. At specific days post infection, the eels were euthanized to measure the bioluminescence radiance using IVIS for the both sides of the body, gills and internal organs. We tested the potential of different concentrations of AngHV-1 UK ORF35 Del strain as a vaccine candidate for the mass vaccination of yellow eels by immersion (primary infection) along with a group of eels exposed to psoralen/UV inactivated AngHV-1 ORF35 Del strain (highest dose) to test the hypothesis of infectivity of the virus, acting like an inactivated vaccine. At 36 days post-exposure, eels were challenged with the AngHV-1 UK Luc strain and analysed it in IVIS at specific time points.
Results
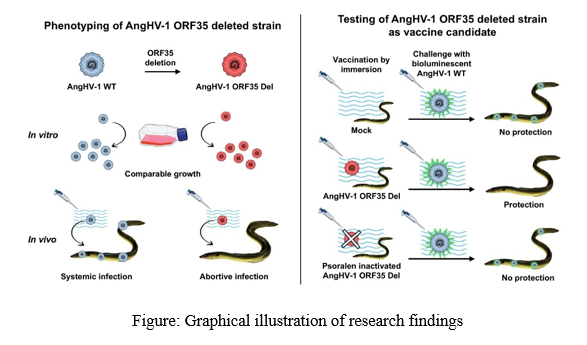
The replication kinetics of ORF35-deleted recombinant strains (UK ORF35 Del and UK Luc ORF35 Del strains) reached high and comparable titers with no significant statistical differences detected with parental strains (UK and UK Luc strains). The UK Luc and the UK Luc ORF35 Del strains expressed both luciferase bioluminescence and copGFP fluorescence, while mCherry fluorescence correlated with ORF35 deletion. Plaque-area assays revealed that the two ORF35 Del strains produced significantly smaller plaques than the wild-type parental strains. AngHV-1 ORF35-deleted strains using this approach demonstrated that ORF35 is not essential for viral replication in cell culture. For IP inoculation in eels, UK Luc ORF35 Del strain expressing either much lower or no bioluminescence signals when compared with UK Luc strain. Similar to IP, immersion infection revealed significant differences between parental and ORF35 Del strains. While intradermal inoculation showed that the fish infected with either the UK Luc or UK Luc ORF35 Del strains showed positive signals at 2 dpi but only in the skin then these signals reduced at later time points in eels infected with UK Luc ORF35 Del. This suggested that the inoculation led to an erratic and abortive infection despite the artificial systemic mode of infection used. In the case of different concentrations of UK ORF35 Del strain as a vaccine candidate, at 6 dpi and at later timepoints, all the eels from mock and Psoralen/UV-inactivated groups expressed infection in the skin, gills and also in the other organs tested (brain, heart, and gut–liver). A significant effect of UK ORF35 Del was observed for all doses tested for the skin and the gills, while a significant effect was observed for the brain, the heart, and the gut–liver at doses of 50,000 pfu/mL or higher. It shows the ability of ORF35 Del strain to confer a protective immunity against a wild-type challenge.
Conclusion
The present study investigated, using recombinant strains and bioluminescent imaging, the effect of AngHV-1 ORF35 deletion on virus replication in vitro, virulence in vivo, and the potential of AngHV-1 ORF35-deleted recombinants as vaccine candidates for the mass vaccination of eels by immersion. The results suggest that the AngHV-1 ORF35 expresses singular and essential properties in vivo compared to its orthologues encoded by CyHV-2 and CyHV-3. This study supports the potential of AngHV-1 ORF35-deleted recombinants as vaccine candidates for the mass vaccination of eels by immersion.