SEX RATIO AND SEXUAL DIMORPHISM IN THREE NATURAL POPULATIONS OF EUROPEAN SEABASS Dicentrarchus labrax EXPOSED TO FOUR THERMAL REGIMES
Introduction
With global warming, the surface temperature of the Mediterranean Sea has increased by 0.4 °C per decade from 1985 to 2006 (Nykjaer, 2009) and is projected to increase further by 2.2 to 3.4 °C until 2080 (IPCC, 2021). The European seabass, Dicentrarchus labrax, is very important for both aquaculture and fisheries in the Mediterranean. One of the big challenges in seabass farming is to improve the sex ratio in favour of the females, which grow faster and mature later. Moreover, seabass has a polygenic sex determination system with environmental influences, mainly driven by temperature (Vandeputte and Piferrer, 2019). Therefore, studying the impact of changing temperature on sex ratio is crucial for seabass production. In this study, we investigated the sex ratio and sexual dimorphism of three natural populations: Atlantic (AT), Western Mediterranean (WM) and Eastern Mediterranean (EM) in different thermal regimes.
Materials and Methods
The production of the three populations was achieved using three different full factorials with 30 sires per population and 7, 14 and 11 dams for the AT, WM and EM populations, respectively. Until 20 days post-hatching (dph), the populations were reared separately at 13°C in 2 replicate tanks per population. From 20 dph, the populations were mixed in equal proportions and placed in four thermal regimes that mimic the temperatures encountered in the Atlantic (rAT, annual average = 13.8 °C, min = 10.4 °C, max = 18 °C), the western Mediterranean Sea (rWM, annual average = 16.6 °C, min = 12.6 °C, max = 22.5 °C), the eastern Mediterranean Sea (rEM, annual average = 21.7 °C, min = 16.8 °C, max = 27.6 °C) and a temperature favouring females applied in aquaculture (rAQUA, 16 °C until 70 dph, 23 °C from 70 to 120 dph and then an eastern Mediterranean temperature regime). At a weight of 10 g, 6230 fish were tagged, weighed and genotyped with the AxiomTM Seabass 57k SNP DlabChip array. To recover the pedigree and population of each fish, parentage assignment was performed by exclusion using a subset of 1000 markers with the APIS R package (Roche et al., 2024), with two allelic mismatches tolerated. Each fish was sexed by macroscopic examination of the gonads at slaughter which occurred between 1 and 3 years of age in different fish groups. Threshold genomic models using the blupf90 suite of programs and the genomic relationship matrix were used to estimate heritabilities and genetic correlations, and to assess genotype-by-environment interaction (GxE) of sex between regimes.
Results
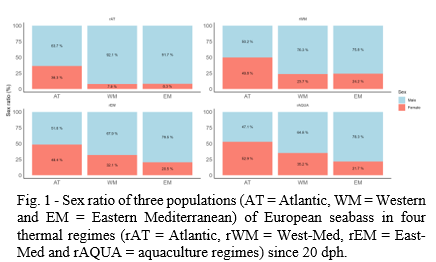
5704 fish were unambiguously assigned to their parents and sexed as a male or a female. A significant interaction was observed between thermal regime and population on sex ratio. For each population, significantly less females were observed in rAT than in the other regimes (P < 1e-04). For the WM population, significantly less females were observed in rWM than in rAQUA (P < 1e-04). In each thermal regime, there were more females in the AT population compared to WM and EM. For the warmer regimes (rEM and rAQUA), all populations were significantly different from each other, and WM had more females than the EM population (Fig. 1). Globally, the sex ratios of the AT population were balanced with an average of 46.9 % of females whereas the ones for Mediterranean populations biased toward males, even more in colder regimes (rAT and rWM).
A 3-way interaction between regime, population and sex was significant for body weight at the time of tagging (≈ 10 g, P = 1.19e-04), meaning that the sex dimorphism differed among populations and thermal regimes. In colder regimes (rAT and rWM), no interaction was found between sex and population and females were 28.5 % heavier than males on average. In the warmer regimes, the interaction between sex and population was significant (P < 8.08e-04). The difference in weight between females and males was stronger for the AT population (> 50 %) compared to Mediterranean populations (33-42 %). The heritability of sex was similar between AT and EM populations, averaging 0.64 ± 0.07. Further analyses will be presented, especially regarding GxE for sex-ratio..
Discussion
The results confirmed that the sex of European seabass is strongly influenced by temperature but also has a strong genetic basis, showing for the first time a population effect with the AT population producing more females. We also showed that the sexual size dimorphism of body weight in favour of females depended on population and thermal regime with higher sexual dimorphism of body weight for the AT population in warmer regimes (rEM and rAQUA). Globally, we decrypted two groups of thermal regimes with one represented by rAT having temperatures below 13 °C until 50 dph and one represented the other thermal regimes. Indeed, the rAT had sex ratio more biased toward males than the other thermal regimes for the three populations and confirmed that a long lasting of cold rearing temperature promote the proportion of males (Vandeputte et al., 2020). Further analyses will describe why AT population expressed higher proportion of females with the results of heritability and genetic correlations but also studying the sex determination QTLs between the populations. This study is part of FishNess project (ANR-21-CE20-0043).
References
IPCC (2021) Climate Change 2021 - The physical science basis. 3949
Nykjaer L (2009) Mediterranean Sea surface warming 1985–2006. Clim Res 39: 11–17
Roche J, Griot R, Allal F, Besson M, Haffray P, Patrice P, Phocas F, Vandeputte M (2024) APIS: an updated parentage assignment software managing triploids induced from diploid parents. G3 GenesGenomesGenetics 14: jkae143
Vandeputte M, Clota F, Sadoul B, Blanc M, Blondeau‐Bidet E, Bégout M, Cousin X, Geffroy B (2020) Low temperature has opposite effects on sex determination in a marine fish at the larval/postlarval and juvenile stages. Ecol Evol 10: 13825–13835
Vandeputte M, Piferrer F (2019) Genetic and Environmental Components of Sex Determination in the European Sea Bass. In H-P Wang, F Piferrer, S-L Chen, Z-G Shen, eds, Sex Control Aquac. John Wiley & Sons, Ltd, Chichester, UK, pp 305–325