ELEVATED VITAMIN LEVELS ENHANCE WOUND HEALING EFFICIENCY IN RAINBOW TROUT
Introduction
Lesions in the skin are a major welfare problem for the fish and a big cause of economic losses for the salmon industry (Takle et al. 2015). Skin lesions, abrasions, and ulcers are the main entrance for pathogens. In these situations, skin wounds must be promptly treated in order to avoid negative impacts on fish (Ceballos et al. 2017 and Costa et al. 2018) . The wound healing process is dependent on factors such as temperature, wound dimensions, stress, and importantly nutrition. However, few studies have investigated nutritional interventions that could potentially improve the regeneration of fish skin (Sveen et al 2019) . The aim of this study was to test the effects of vitamin inclusion level on wound healing. To this end a skin wound healing model was created in salmonids to evaluate the molecular and biochemical processes involved in the different phases of regeneration.
Material and Method
The trial was performed at the Research Centre for Animal Nutrition and Health (DSM Nutritional Products France). Eighty rainbow trout (Oncorhynchus mykiss; initial body weight 63g) were randomly allocated to two different experimental groups. The fish were fed 84 days with basal diet mimicking a Norwegian commercial recipe (control) or an Optimum Vitamin Nutrition (OVN) diet (Liu et al 2022) that differed from the control in higher vitamin level inclusions. Each treatment was given to two replicate groups of 20 fish. Animals from the 2 diets were wounded after 28 days of feeding with the experimental diets corresponding to day 0 of the experiment. The fish were wounded with a punch biopsy tool, transversing the full thickness of the skin barrier on the left dorsal flank of the fish. The principal advantage of this model is that a deep wound involves both dermal and epidermal repair processes. The fish continued to be fed with the experimental feed until the end of the trial. Growth parameters, blood biochemistry, wound size, and the expression of genes representative of the different phases of the wound healing process were monitored at day 0, 1, 3, 7, 14, 21, 28, 35, 49 and 56 days post wounded (dpw).
Results
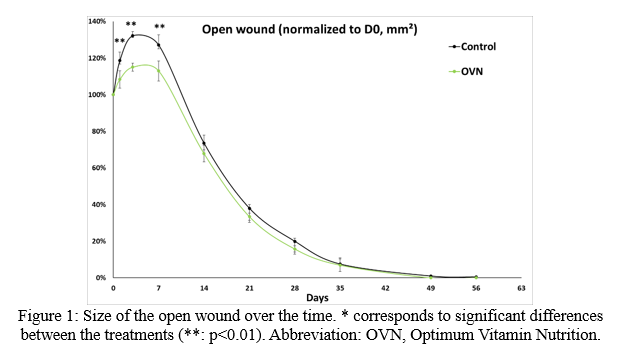
As expected, strong modulations over time were observed for all the blood parameters, except for phosphorus (p=0.0870) and sodium (p=0.1035). An increase in the size of the wounds on 1dpw, 3dpw, and 7dpw was observed. The return of the initial size of the wounds occurred for the two treatments between 7dpw and 14dpw . With OVN diet a complete closure of all the wounds occurred 7 days before those of fish fed with control diet. After normalization of the size of the wounds to the 0dpw size for each fish, statistical differences were observed between the two treatments at 1dpw (p=0.0028), 3dpw (p=0.0014) and, 7dpw (p=0.0072). From 14dpw to the end of the trial, no differences were observed between the treatments (p>0.05) (Figure 1).
We were able to identify the progression of wound healing at the transcriptomic level. The re-epithelialization genes followed an expected pattern, but maximum expression was delayed. For MMP13 we observed maximum expression at 14dpw for control diet and 7dpw for OVN diet. Maximum expression of chemokine 4 ligands was observed at 3dpw for the two diets. This may reflect the fact that immune cells are less necessary at the very beginning of the wound healing process in our model. The inflammation step was reduced in this trial. The expression of serum amyloid A appeared early, at 3dpw for both diets. In addition, IL-1B gene expression levels were flat. Granulation tissue genes are also affected in our model. Instead of a continuously decreasing expression expected , we observed a maximum of expression at 14 days with the two diets. This could indicate a lack of activation of the gene at the very beginning of the wound . The genes involved in tissue remodeling (Col1a1a and Hyaluronisase-2- precursor) showed an expected profile, similar between the two diets.
The genes involved throughout the wound healing process (Cytochrome P450 4F3 and Epigen precursor) showed also an expected expression profile, reflecting the success of the wound healing process. Interestingly, t he level of CD4 expression was lower with control diet than with OVN diet.
Conclusion
The differences between the inclusion levels of vitamins in diets had an impact in the wound healing processes. W e were able to observe differences not only macroscopically but also at transcriptomic level supporting that vitamins play a key in wound healing. Then, the reduction of vitamin inclusion levels in salmonid diets should be done carefully.
Bibiography
Ceballos-Francisco D, Cordero H, Guardiola F, Cuesta A, Esteban M.A, Healing and mucosal immunity in the skin of experimentally wounded gilthead seabream (Sparus aurata L), Fish Shellfish Immunol. 71 (2017) 210–219.
Costa R.A, D.M. Power D.M , Skin and scale regeneration after mechanical damage in a teleost, Mol. Immunol. 95 (2018) 73–82
Liu A, Thi Ha To V P, Santigosa E, Dumas A, Hernandez J M, Vitamin nutrition in salmonid aquaculture: From avoiding deficiencies to enhancing functionalities, Aquaculture,Volume 561,2022,738654,
Richardson R er al. Adult zebra fish as a model system for cutaneous wound healing research. The journal of investivative dermatology 133, 1655-1665 (2013).
Sveen LR, Timmerhaus G, Krasnov A, Takle H, Handeland S, Ytteborg E. Wound healing in post-smolt Atlantic salmon (Salmo salar L.). Sci Rep. 2019 Mar 5;9(1):3565.
Takle H , et al. In Norwegian Sårptoblematikk og hudhelse i laks- or regnbueørrettoppdrett. English abstract; Wounds and skin welfare in Atlantic salmon and Rainbow trout )2015)