THE LASTING EFFECTS OF EMBRYONIC TEMPERATURE ON ATLANTIC SALMON IMMUNITY TO Yersinia ruckeri
Introduction
In both Scottish and Norwegian aquaculture, around one fifth of Atlantic salmon (Salmo salar ) are lost during the seawater phase of production due to a wide range of pathogens, underscoring the need for more robust and resilient fish. One possible contributor to low robustness is suboptimal temperature during embryogenesis. In many species, including Atlantic salmon, exposure to environmental stimuli such as temperature during embryogenesis can induce long-lasting effects on physiology and metabolism, a phenomenon known as metabolic programming (Ziqiang et al., 2019). Notably, embryonic temperature in salmon is positively associated with growth rate during early development (Gorodilov, 1996).
In the COOLFISH project, we hypothesised that relatively higher embryonic temperature regimes sometimes used in salmon aquaculture may lead to biased resource allocation towards growth, with negative effects on immune system development . In support of this hypothesis, a r ecent study demonstrated embryonic temperature-dependent differences in Atlantic salmon mortality following Y. ruckeri exposure at the alevin stage, indicative of higher robustness at 4 oC than 8 oC (Malik et al., 2025) . Currently, it remains unclear whether the impact of embryonic temperature on fish robustness is maintained until later life stages due to long-term changes in immune function. In this study, we investigate how changing temperature solely from fertilisation to the eyed stage of Atlantic salmon embryogenesis influences the functional immune response to Yersinia ruckeri infection using head kidney (i.e., primary immune tissue) samples at the parr stage using a transcriptomic approach .
Methods
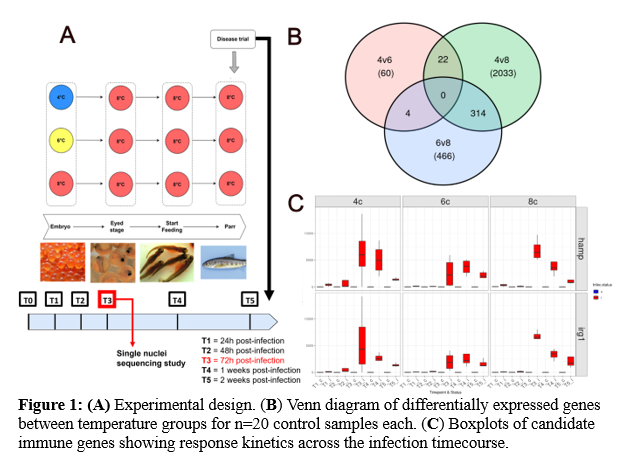
Atlantic salmon embryos were reared under three temperature regimes (4 °C, 6 °C, and 8 °C) from fertilisation to the eyed stage, after which all groups were maintained at 8 °C until start-feeding, followed by 10 °C until sampling (Figure 1A) . Approximately nine months after the end of the embryonic temperature treatment, head kidney tissue was sampled from 120 parr-stage salmon (n = 4 per treatment group, for both control and Yersinia ruckeri-infected fish) across five timepoints (T): T1 (24 h), T2 (48 h), T3 (72 h), T4 (1 week), and T5 (2 weeks) post-infection (Figure 1A). Total RNA was extracted using a Promega Maxwell instrument before its purity and integrity was confirmed. mRNA-seq libraries were produced and sequenced on an Illumina platform by Novogene . A lignment and quantification of reads was carried out through the NF-core RNA-seq pipeline (v3.14) using STAR and RSEM respectively . Using edgeR (v4.4.2) a generalised linear model was constructed to assess differences in infection dynamics across temperature groups, track the immune response over time, and explore interactions between these factors. Additionally, self-organising map (SOM) clustering was applied to identify key gene clusters responsive to infection and temperature. Differential expression tests were conducted for all pairwise temperature comparisons (4 °C vs. 6 °C, 4 °C vs. 8 °C, and 6 °C vs. 8 °C) , across all timepoints, with Gene Ontology (GO) enrichment analysis used to interpret the biological functions of significantly differentially expressed genes.
Results and Discussion
Pairwise differential gene expression analysis of head kidney control samples (n=20 per temperature group, drawn from T1-T5 control samples) revealed 2,067 genes to be constitutively differentially expressed between temperature groups (p-adj < 0.05, logFC>0.5) (Fig. 1.B ). The 8 °C group was a clear outlier, showing a much higher number of differentially expressed genes compared to both the 6 °C (466 genes) and 4 °C (2033 genes) groups. In contrast, the 4 °C and 6 °C groups were more similar, with only 60 differentially expressed genes. Cumulative mortality over the course of the Yersinia ruckeri challenge differed by embryonic temperature group, with the 4 °C group showing the lowest mortality (~22%), followed by the 6 °C (~31%) and 8 °C (~35%) groups. 3,935 genes were differentially expressed in the head kidney in response to Yersinia ruckeri infection across all timepoints and temperature groups (p-adj <0.01, logFC >1) . Of these, 2, 475 genes displayed temperature group specific expression patterns. Temporal profiling of key immune marker genes revealed T3 to be when the peak inflammatory response occurred in all temperature groups (Figure 1C) . However earlier upregulation of numerous immune genes was observed in the 4 °C group, suggesting a more rapid immune activation. This pattern was further supported by enrichment of immune-related GO terms specific to early timepoints (T1 and T2) in the 4 °C group.
Biological interpretation of differential gene expression among temperature groups is ongoing. However, our findings provide clear initial evidence that embryonic temperature exerts long-lasting effects on head kidney gene expression, persisting to the parr stage and shaping the host immune response to bacterial infection. We have also generated head kidney single-nucleus RNA-seq data from timepoint three to gain insights into the cell-type specific nature of these temperature-induced effects during the peak inflammatory response.
The Norwegian Research Council funded this study (Grant number: 325571)
References
Gorodilov, Y. N. (1996). Description of the early ontogeny of the Atlantic salmon, Salmo salar, with a novel system of interval (state) identification. Environmental Biology of Fishes, 47(2), 109–127.
Malik, M. S., Rebl, A., Burgerhout, E., & Lazado, C. C. (2025). Embryonic Temperature Influences the Mucosal Responses of Atlantic Salmon Alevins to a Bacterial Challenge. Marine Biotechnology, 27(1), 1-12.
Ziqiang , Z., Fang, C., Xiaozhong , L. (2019). Epigenetic Programming and Fetal Metabolic Programming. Frontiers in Endocrinology, 10.